توضیحات
اسیدهای چرب جیره از طریق بیوهیدروژناسیون تغییرات ساختاری قابل توجهی می کنند زمانیکه از شکمبه عبور می کنند و برای جذب به روده ها منتقل می شوند. بخش قابل توجهی از تولید چربی شیر، که عامل اصلی درآمد شیر است، توسط این مسیرهای بیوهیدروژناسیون هدایت می شود. تغییر ترکیبات مواد مغذی خوراک که یا به طور ناخواسته یا به دلیل تنوع مواد مغذی در مواد تشکیل دهنده خوراک ایجاد می شوند، می توانند مسیرهای بیوهیدروژناسیون را تغییر دهند و باعث تغییر در خروج چربی های فعال زیستی از شکمبه شوند که بر تولید چربی شیر تأثیر منفی می گذارند. بنابراین، شناسایی عواملی که باعث انتقال بیوهیدروژناسیون اسیدهای چرب در شکمبه از "چربی شیر دوستانه" به "چربی شیر غیر دوستانه" می شوند از اهمیت بالایی برخوردار است.
هدف این مقاله ارائه مکانیزم محرک احتمالی است که جمعیت میکروبی شکمبه را برای تغییر مسیرهای بیوهیدروژناسیون به سمت نامطلوبی برای تولید چربی شیر آغاز می کند. بسیاری از شواهد مستقیم در مورد عوامل ایجاد کننده از مطالعات اخیر در مورد باکتری های جدا شده شکمبه، کشت شکمبه در آزمایشگاه و داده های گاو به دست امده است. داده ها در سراسر این مطالعات نشان می دهند هنگامی که اسیدهای چرب جیره، هم از جیره اصلی و هم از مکمل چربی، به سطحی برسند که باعث ایجاد اثرات ضد باکتریایی در شکمبه شوند، نتیجه تغییر از بیوهیدروژناسیون طبیعی به یک مسیر جایگزین است. مسیر جایگزین تولید واسطه های فعال زیستی لیپیدی می کند که چربی شیر را کاهش می دهد. داده های خلاصه شده در زیر همچنین نشان می دهند که نوع و غلظت اسید چرب مورد نیاز برای رسیدن به اثرات ضد باکتریایی تحت تأثیر سایر ملاحظات مربوط به مواد مغذی در جیره است.
اسیدهای چرب چیست؟
قبل از شروع بحث در مورد سرنوشت اسیدهای چرب هنگام عبور از شکمبه، توضیح مختصری در مورد تعریف اسیدهای چرب می دهیم. به بیان ساده، اسیدهای چرب مهمترین عناصر سازنده چربی هستند همانطور که اسیدهای آمینه عناصر سازنده پروتئین هستند. اسیدهای آمینه به همراه پیوندهای پپتیدی در طول های مختلف به هم متصل می شوند و از دی پپتیدها (2 اسید آمینه) تا پلی پپتیدها (10 < اسید آمینه) تشکیل می شوند. برخلاف پروتئین، چربی ها از بیش از سه اسید چرب تشکیل نمی شوند که به عنوان پیوست روی ستون فقرات گلیسرول در کنار هم قرار گرفته اند. چربی ها و روغن ها در درجه اول از سه اسید چرب متصل به گلیسرول تشکیل می شوند که به آنها تری گلیسیرید (یا به عبارت صحیح تر تری اسیل گلیسرول) گفته می شود. لیپیدهای علوفه ای عمدتاً حاوی گالاکتولیپیدها هستند، جایی که ستون فقرات گلیسرول دارای دو اسید چرب متصل به همراه یک مولکول قند است.
اسیدهای چرب و نه ستون فقرات گلیسرول، مزایایی برای عملکرد دام از جمله انرژی زیاد، فواید بافتی و اثرات شکمبه ای را فراهم می کنند. به همین دلیل، مهم است که درک اساسی از تفاوت بین اسیدهای چرب داشته باشیم. اسیدهای چرب زنجیره های کربن هستند که به یک گروه اسیدی یا همان گروه کربوکسیل ختم می شوند همانطور که در بیوشیمی اشاره شده است. یک نمونه از اسیدهای چرب رایج، اسید استئاریک با 18 کربن و بدون پیوند مضاعف است (شکل 1). اسید استئاریک در روغن های گیاهی کم است، اما در چربی های حیوانی، به ویژه در چربی های حاصل از گونه های نشخوارکننده مانند پیه گاو، مقادیر بیشتری وجود دارد.
اسیدهای اولئیک، لینولئیک و لینولنیک نمونه هایی از اسیدهای چرب غیراشباع حاوی یک یا چند پیوند مضاعف هستند (شکل 2). اسید اولئیک یک پیوند مضاعف بین کربن های 9 و 10 دارد و از آن به عنوان اسید چرب غیراشباع مونو نام برده می شود. اسید لینولئیک نوعی اسید چرب غیراشباع است که حاوی دو پیوند مضاعف بین کربن های 9 و 10 و بین کربن های 12 و 13 است. اسید اولئیک اسید چرب غالب در چربی های حیوانی و برخی روغن های گیاهی مانند روغن کانولا است. اسید لینولئیک اسید چرب غالب در بسیاری از روغن های گیاهی از جمله روغن پنبه دانه، روغن سویا و روغن ذرت است. اسید لینولنیک، با سه پیوند دوگانه، اسید چرب اولیه در بیشتر گونه های مرتعی و در روغن بذر کتان است.
ورودی اسیدهای چرب به شکمبه
میزان مصرف روزانه رایج اسیدهای چرب غیراشباع برای جیره ها با و بدون مکمل چربی در جدول 1 نشان داده شده است. غلظت کل اسیدهای چرب در جیره ها می تواند از کمتر از 20 میلی گرم در گرم ماده خشک برای مواد پایه تا بیش از 80 میلی گرم هنگام اضافه شدن چربی باشد. اسید لینولئیک اسید چرب غیراشباع غالب است که در بیشتر موارد به مقدار بیش از 700 گرم در روز در مطالعات منتشر شده مصرف می شود (جدول 1)، یا حتی بیش از 1000 گرم در روز در شرایط مزرعه (Chase, 2019).
غلظت اسیدهای چرب در محتوای شکمبه نشان دهنده غلظت و تنوع آنها در خوراک است. به عنوان مثال با استفاده از نتایج سه مطالعه منتشر شده (جدول 2)، اسیدهای چرب در محتوای شکمبه از کمتر از 10 میلی گرم در هر گرم ماده خشک برای جیره حاوی 50% یونجه برموداگراس (Bateman and Jenkins, 1998) تا 29 میلی گرم در گرم ماده خشک برای جیره یونجه و سیلاژ ذرت (Loor et al., 2002)، و زیر 50 میلی گرم در گرم ماده خشک برای جیره با روغن های گیاهی مکمل متفاوت بودند. جالب توجه است که گاوهایی که از مراتع شبدر استفاده می کنند، دارای غلظت اسید چرب شکمبه نزدیک به سطوح مشاهده شده برای جیره های با مکمل چربی هستند. مفهوم حفظ اسیدهای چرب در محتوای شکمبه این است که بسیاری از گونه های میکروبی به غلظت های بالای اسید چرب حساس هستند و با کاهش رشد و فعالیت متابولیکی پاسخ می دهند. به طور خاص، فعالیت ضد باکتری برای اسیدهای چرب غیراشباع بیشتر است و از ویژگی های اسیدهای چرب اشباع نیست.
باکتری های شکمبه ای یک مکانیسم محافظتی ذاتی دارند که برای کاهش غلظت اسید چرب غیراشباع در شکمبه و کاهش احتمال فعالیت ضد باکتری طراحی شده است. از این مکانیسم محافظتی به عنوان بیوهیدروژناسیون یاد می شود، جایی که اسیدهای چرب غیراشباع به صورت آنزیمی به اسیدهای چرب اشباع تبدیل می شوند (Jenkins et al., 2008). کارآیی بیوهیدروژناسیون را می توان از نتایج سه مطالعه در جدول 2 مشاهده کرد که در آن غلظت اسید چرب غیراشباع شکمبه در مقایسه با غلظت اسید چرب کل خوراک یا شکمبه بسیار کمتر و کم تنوع تر است. بیوهیدروژناسیون، در حالی که به جمعیت میکروبی در کنترل اثرات ضد باکتریایی اسیدهای چرب غیراشباع کمک می کند، ماهیت خروج اسید چرب از شکمبه را نیز در مقایسه با ورودی آن از خوراک بسیار تغییر می دهد. این فرآیند تبدیل اسیدهای چرب پیامدهای مثبت و منفی بر تولید دام و قابل قبول بودن محصولات غذایی حیوانی دارد.
تغییر غلظت اسیدهای چرب شکمبه در طول زمان
مسیرهای بیوهیدروژناسیون بسیار پیچیده هستند و طیف گسترده ای از واسطه ها را تولید می کنند. سه اسید چرب اشباع نشده اصلی که مصرف می شوند (اسیدهای اولئیک، لینولئیک و لینولنیک) همگی تحت تغییرات آنزیمی قرار می گیرند که باعث ایجاد تعداد زیادی از واسطه های منحصر به فرد می شود. به عنوان مثال، مسیرها و واسطه های بیوهیدروژناسیون اسید لینولئیک و لینولنیک به اسید استئاریک در شکل 3 نشان داده شده است (Ferlay et al., 2017). با افزایش دانش در مورد مسیرهای بیوهیدروژناسیون، هویت واسطه ها گسترش می یابد. ورود فقط 3 اسید چرب غیراشباع (اولئیک، لینولئیک و لینولنیک اسید) به عنوان مواد اولیه در شکمبه منجر به تولید ده ها، اگر نه صدها ایزومر اسید چرب پیچیده در خروج از شکمبه می شود. با این حال، این پیچیدگی بیوهیدروژناسیون در اکثر بحث های مربوط به بیوهیدروژناسیون تا حد زیادی نادیده گرفته می شود. بیشتر توجهات معطوف به نسخه ساده تری از بیوهیدروژناسیون اسید لینولئیک است (شکل 4) که فقط بر چند ماده میانی تأکید دارد که قبلاً نشان داده شده بودند که از تولید چربی در غده پستانی جلوگیری می کنند.
خیلی خلاصه، بیوهیدروژناسیون اسید لینولئیک در شکمبه با تبدیل آن به اسید لینولئیک مزدوج (CLA) آغاز می شود. در این مرحله اولیه، تعداد پیوندهای مضاعف ثابت باقی می ماند اما یکی از پیوندهای مضاعف توسط آنزیم های میکروبی به موقعیت جدید منتقل می شود. به طور معمول، پیوندهای مضاعف در اسید لینولئیک توسط دو پیوند منفرد از هم جدا می شوند، اما در CLA، پیوندهای مضاعف فقط با یک پیوند واحد جدا می شوند. انواع بسیاری از CLA در شکمبه گاوهای شیری تولید می شوند، اما یک CLA متداول که از بیوهیدروژناسیون اسید لینولئیک تولید می شود، cis-9 ، trans-11 C18: 2 است.
با پیشرفت بیوهیدروژناسیون، پیوندهای مضاعف در واسطه های CLA به اسیدهای چرب ترانس که فقط یک پیوند مضاعف دارند هیدروژنه می شوند. مرحله نهایی هیدروژناسیون توسط میکروب های شکمبه آخرین پیوند مضاعف را حذف می کند و اسید استئاریک را به عنوان محصول نهایی تولید می کند. پیوندهای مضاعف ترانس فقط از نظر قرارگیری هیدروژن ها با پیوندهای مضاعف سیس فرق دارند. هیدروژن ها در طرف مقابل پیوند مضاعف اسیدهای چرب ترانس، اما در یک طرف پیوند مضاعف اسیدهای چرب سیس قرار دارند. اگرچه تفاوت ساختار بین اسیدهای چرب ترانس و سیس اندک به نظر می رسد، اما تفاوت چشمگیری در خصوصیات فیزیکی و متابولیکی آنها ایجاد می کند. مسیر ترانس 11(t11) بیوهیدروژناسیون اسید لینولئیک در شکل 4 شامل واسطه هایی از جمله cis-9, trans-11 CLA است که اثبات شده تأثیر اندکی بر چربی شیر دارد. مسیر ترانس -10 (t10) شامل واسطه هایی از جمله trans-10, cis-12 CLA است که ثابت شده تولید چربی شیر را کاهش می دهد (Baumgard et al., 2000).
یک آزمایش مزرعه ای از افزایش اسیدهای چرب غیراشباع در محتوای شکمبه در داده های Baldin et al. (2018) مشاهده شد. آنها بیشترین غلظت اسیدهای چرب غیراشباع در محتوای شکمبه گاوها را در عرض 5 دقیقه با دوز داخل رحمی 200 گرم روغن غیراشباع گزارش کردند. غلظت شکمبه ای اسیدهای چرب غیراشباع پس از 4 ساعت از تزریق به مقادیر اصلی بازگشت. این نشان می دهد که گاوهایی که تحت شرایط مزرعه ای تغذیه می شوند بلافاصله پس از تغذیه جهشی در غلظت اسید چرب غیراشباع در شکمبه را تجربه می کنند که ممکن است برای ایجاد اثرات ضد باکتریایی در شکمبه کافی باشد. دامنه افزایش احتمالاً تابعی از اسیدهای چرب ورودی از جیره، درصد غیراشباع بودن اسیدهای چرب جیره، دفعات تغذیه و میزان مصرف خوراک است.
اثرات ضد باکتریایی اسیدهای چرب غیراشباع
مطالعات گسترده ای در مورد توصیف فعالیت ضد باکتریایی اسیدهای چرب مختلف وجود دارد (Desbois and Smith, 2010). دو عاملی که بر فعالیت ضد باکتریایی چربی ها تأثیر می گذارند، ساختار و غلظت اسیدهای چرب است. اسیدهای چرب آزاد به طور کلی تخمیر را بیش از تری گلیسیرید مختل می کند و فعالیت ضد باکتریایی اسیدهای چرب آزاد را می توان با افزایش تعداد پیوندهای مضاعف افزایش داد (Desbois and Smith, 2010). رشد برخی از گونه های باکتریایی با غلظت کم اسیدهای چرب تحریک می شود، اما در غلظت های بالاتر مهار می شود (Maczulak et al., 1981). در تلاش برای پیش بینی تغییرات تخمیر شکمبه ناشی از چربی جیره، اغلب فرض بر این است که میزان چربی فقط با مکمل چربی تأمین می شود و غلظت اسیدهای چرب آزاد (FFA) کم است. هر دو فرض می تواند اشتباه باشد. اسیدهای چرب حاصل از TMR و علوفه می تواند به میزان قابل توجهی در میزان کل چربی شکمبه تاثیر داشته باشد، به عنوان مثال هنگامی که حیوانات از مرتع نابالغ تغذیه می کنند. همچنین، غلظت اسیدهای چرب آزاد ممکن است در بعضی از مواد خوراکی مانند پنبه دانه کامل که در شرایط گرم و مرطوب نگهداری می شود، افزایش یابد (Cooke et al., 2007)، یا در علوفه های حاصل از تجزیه هیدرولیتیک چربی های استری شده در حین تولید علوفه (Yang and Fujita, 1997).
مکانیسم های اثرات ضد باکتریایی اسیدهای چرب در درجه اول به نفوذ آنها به غشای سلول باکتریایی وابسته است. جزئیات فرایندهای مکانیکی (شکل 6) را می توان بر اساس رابطه بین سه جنبه زیر طبقه بندی کرد: 1- افزایش نفوذپذیری غشا و تخریب سلول، 2 - اختلال در زنجیره انتقال الکترون و فسفوریلاسیون اکسیداتیو جدا شده، و 3- مهار فعالیت های آنزیمی غشا و جذب مواد مغذی (Yoon et al., 2018). برای میکروب های بی هوازی که در شکمبه زندگی می کنند، اسیدهای چرب از طریق بی نظمی در غشای خارجی سیتوپلاسمی دارای خواص ضد باکتری هستند که می تواند منجر به افزایش نفوذپذیری غشا و حتی تخریب سلول، مهار فعالیت آنزیمی غشا و اختلال در جذب مواد مغذی شود.
چندین ویژگی از اثرات ضد باکتریایی در شکمبه مستقیماً بر مسیرهای t10 در مقابل t11 بیوهیدروژناسیون و تأثیر نهایی بر چربی شیر تأثیر می گذارد.
1- یک عامل مهم این است که به نظر می رسد اسیدهای چرب اثرات ضد باکتریایی سریعی دارند. Maia et al. (2010) کاهش بیشتر از 96 درصدی فعالیت متابولیکی در باکتری B. fibrisolvens طی 20 دقیقه پس از افزودن 2/0 میلی گرم در میلی لیتر اسید لینولئیک به محیط کشت را گزارش کردند. یک آزمایش در محیط کشت پیوسته که اخیرا در دانشگاه کلمسونClemson انجام شد، بررسی کرد که روغن سویا چگونه سریعا مسیرهای بیوهیدروژناسیون را از مسیر طبیعی و غالب t11 به مسیر فرعی t10 منتقل می کند. محیط کشت حاوی یک جیره پایه برای 4 روز با واسطه های t10-18:1 و t11-18:1که دقیقا قبل از تغذیه صبحگاهی (8 صبح) انالیز می شدند و سپس مجددا در 2 تا 4 ساعت بعد از تغذیه صبح. در روز 5 محیط های کشت به طور ناگهانی به رژیم غذایی حاوی 3% روغن سویا تغییر یافتند و نمونه ها در هر ساعت و به مدت 12 ساعت انالیز شدند. نتایج (شکل 5) افزایش مسیر t10 را پس از افزودن روغن سویا به محیط های کشت در عرض چند ساعت نشان داد. این می تواند به معنای آن باشد که غلظت اسید چرب غیراشباع نیاز نیست که همیشه در سطح بالا باشد تا اثرات ضد باکتریایی ایجاد کند. شاید فقط اوج گذرا در غلظت اسیدهای چرب غیراشباع شکمبه که بلافاصله پس از تغذیه اتفاق می افتد برای ایجاد اثرات ضد باکتری کافی باشد.
2- دومین نکته مهم فعالیت ضد باکتری در شکمبه این است که همه گونه های باکتریایی به یک اندازه حساس نیستند. اختلال در یکپارچگی غشا به دنبال افزودن اسید لینولئیک به محیط کشت تفاوت گسترده ای در 17 گونه باکتریایی شکمبه داشت(Maia et al., 2007) . به طور کلی، گونه های باکتریایی که مسیر t11 بیوهیدروژناسیون را دنبال می کنند اختلال عملکرد غشا را بیشتر نشان می دهند، از جمله بالای 50% اختلال برایButyrivibrio spp. و بالای90% اختلال در.Pseudobutyrivibrio. غشای M. elsdenii که مسیر t10 بیوهیدروژناسیون اسید لینولئیک را دنبال می کند کمتر از 5 درصد با همان دوز اسید لینولئیک مختل شد. بنابراین، غلظت اسیدهای چرب بالاتر از آستانه ضد باکتریایی بسته به گونه های باکتریایی باعث صدمه انتخابی در شکمبه می شوند و آسیب کمتری برای میکروارگانیسم های t10 دیده می شود.
3- همه اسیدهای چرب غیراشباع تمایل مساوی به ایجاد اثرات ضد باکتریایی در همان غلظت ندارند. به عنوان مثال، اثرات مهاری نسبی اسیدهای چرب منفرد در رشد B. fibrisolvens به ترتیب لینولنیک> لینولئیک> اولئیک> استئاریک بودند (Maia et al. 2010). روند کلی مهار بیشتر با افزایش تعداد پیوندهای مضاعف در زنجیره آسیل بود. روندهای مشابه در مطالعات مزرعه ای گزارش شده است. Dorea and Armentano (2017) گزارش دادند که تغذیه اسیدهای چرب اشباع مانند اسید پالمیتیک به گاوها، اسیدهای چرب شیر را به طور عمده با افزایش تولید C16 افزایش می دهد. با این حال، تغذیه اسیدهای چرب غیراشباع با مهار ترشح اسیدهای چرب شیر کوتاه تر از C18 باعث کاهش کل اسید چرب شیر می شود، که اسید لینولئیک بازدارندگی بیشتری از اولئیک دارد.
4- چهارمین و شاید مهمترین خاصیت اثرات ضد باکتریایی این است که آستانه ایجاد تغییر در مسیر بیوهیدروژناسیون توسط شرایط محیطی و شیمیایی شکمبه اصلاح می شود. اگر آستانه غلظت اسیدهای چرب غیراشباع در محتویات شکمبه ثابت بود، توصیه های تغذیه برای چربی بسیار ساده تر می شد. در عوض، نشان داده شده است که pH کم و تجمع اسید لاکتیک اثرات ضد باکتریایی اسیدهای چرب غیراشباع را برجسته می کنند، به طور خاص میکروارگانیسم های t11 را هدف قرار می دهد .(Maia et al., 2010) هر دوی این شرایط، مقادیر بالای نشاسته را به عنوان عامل منفی تولید چربی شیر در نظر می گیرند.
نتیجه گیری
با استفاده از کلیدواژه ضد باکتری توصیف شده در این مقاله، می توان دنباله ای از وقایع را پیشنهاد کرد که به موجب آن مسیرهای بیوهیدروژناسیون را تغییر می دهند و شکمبه را از حالت چربی شیر دوستانه به چربی شیر غیر دوستانه منتقل می کند. مرحله اولیه غلظت اسیدهای چرب غیراشباع در شکمبه است که از آستانه اثرات ضد باکتریایی فراتر رود. این می تواند به یکی از دو روش اتفاق بیفتد. 1) افزایش غلظت اسیدهای چرب غیراشباع در جیره که می تواند ناشی از تغییر اسیدهای چرب پایه یا چربی مکمل به جیره باشد یا 2) کاهش آستانه ضد میکروبی. آستانه تحمل میکروارگانیسم های شکمبه با افزایش نشاسته، کاهش pH شکمبه، افزایش لاکتات یا ترکیبی از آنها کاهش می یابد. پس از رسیدن به آستانه ضد باکتری، میکروارگانیسم های t11 در عرض چند ساعت با خاموش کردن فعالیت متابولیکی شامل نرخ بیوهیدروژناسیون پاسخ می دهند. این موضوع جریان اسید لینولئیک را از طریق مسیر طبیعی بیوهیدروژناسیون t11 کاهش می دهد. در نتیجه، از آنجا که میکروارگانیسم های t10 نسبت به اثرات ضد باکتری حساسیت کمتری دارند، اسید لینولئیک بیشتری از طریق مسیر جایگزین t10 برای بیوهیدروژناسیون موجود است. با هر بار تغذیه بعدی با همان جیره، تجمع (به طور خاص ترانس10 سیس12) در شکمبه ادامه می یابد تا یک جریان ثابت CLA به غده پستانی ادامه یابد که تولید اسیدهای چرب de novo مهار می شود.
برخی از گله های پرتولید بیش از 1000 گرم اسیدهای چرب غیراشباع در روز مصرف می کنند اما هنوز هم چربی شیر را در حدود 4% حفظ می کنند (Chase, L. E., 2019). گله های دیگر کاهش چربی شیر را با کمتر از 500 گرم اسیدهای چرب غیراشباع تجربه می کنند.McCarthy et al. (2018) نتوانستند رابطه معنی داری بین تولید چربی شیر و مصرف اسیدهای چرب غیراشباع در 79 گله در شمال شرقی و غرب امریکا کشف کنند. این تنوع به وضوح نشان می دهد که مصرف اسیدهای چرب غیراشباع به تنهایی پیش بینی خوبی برای چربی شیر نیست. مدل های پیش بینی کننده چربی شیر باید شامل کلیه عواملی باشند که شناخته شده اند و تأثیرات ضد باکتریایی آنها را تحت تأثیر قرار می دهند از جمله مقدار و نوع نشاسته، pH شکمبه (فیبر موثر، نوع و مقدار بافرها، مخلوط کردن TMR و غیره) و نرخ ازادسازی اسیدهای چرب از ساختار گیاه.
جدول 1. متوسط مصرف اسیدهای چرب غیراشباع عمده توسط گاوهای شیری تغذیه شده با TMR بدون و با چربی مکمل به طور متوسط در پنج مطالعه
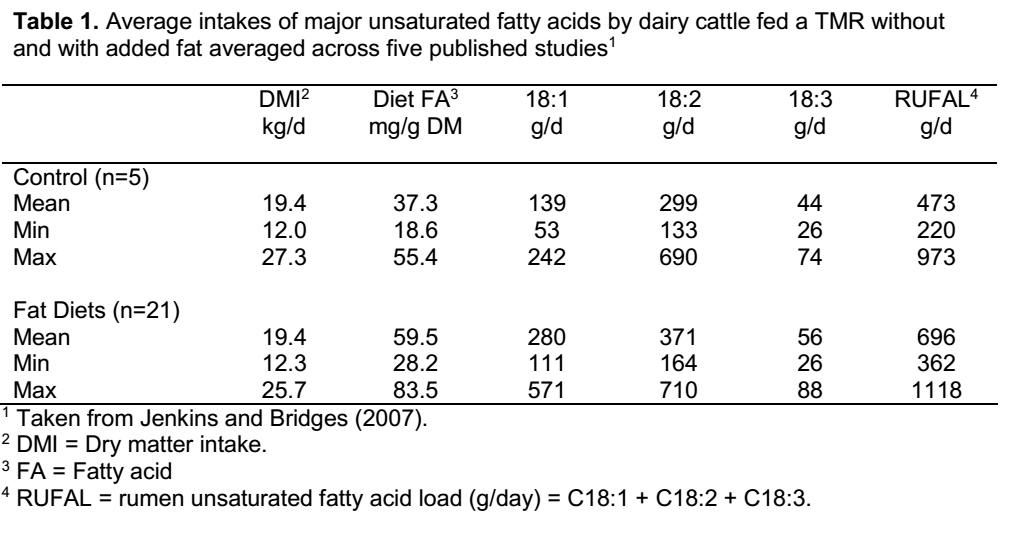
جدول 2. غلظت اسیدهای چرب (میلی گرم در گرم ماده خشک) گزارش شده در نمونه های خوراک و شکمبه از سه مطالعه که گاوها با یک جیره پایه با و بدون روغن مکمل تغذیه می شوند
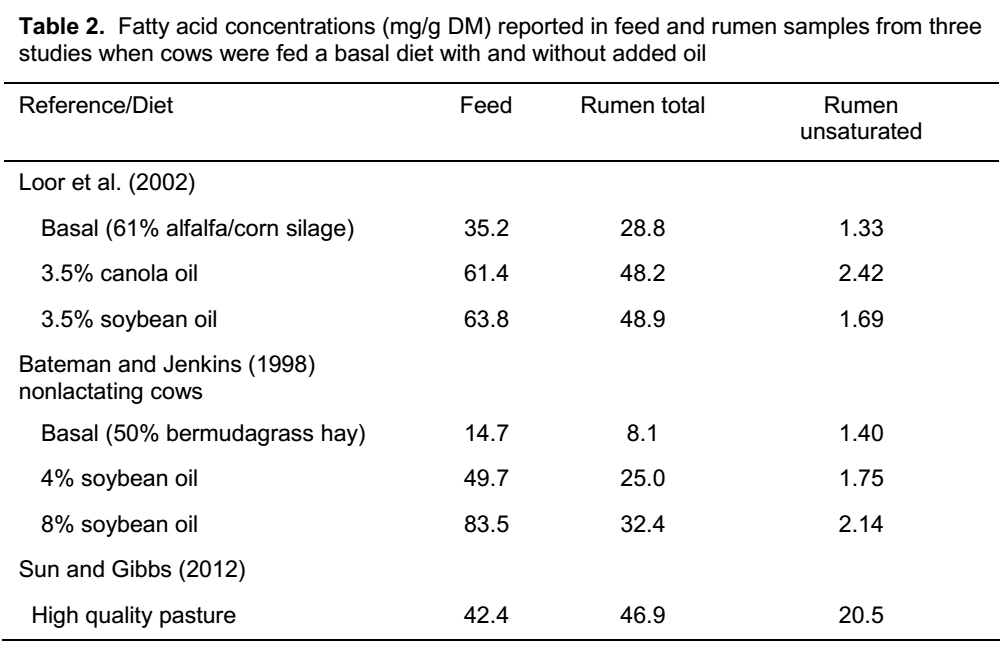
شکل 1. ساختار اسید استئاریک؛ یک اسید چرب اشباع زنجیره بلند
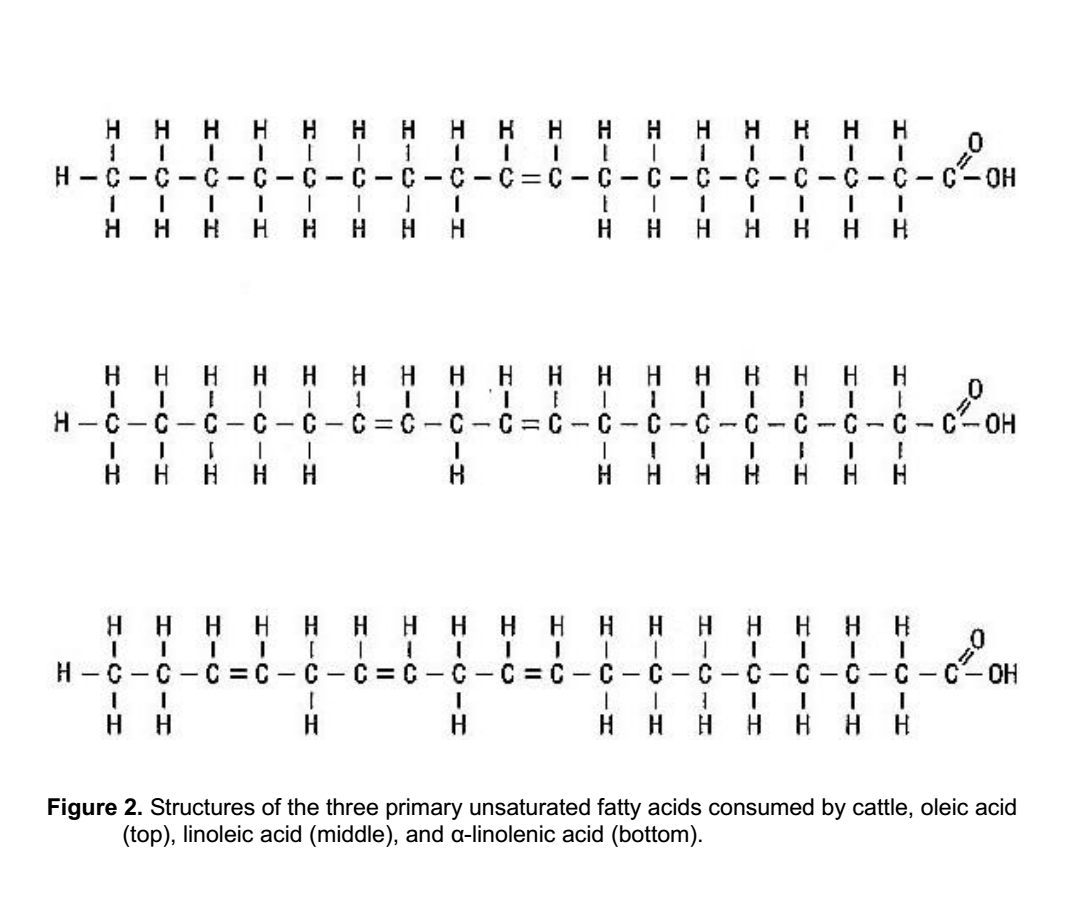
شکل 2. ساختارهای سه اسید چرب غیراشباع اولیه که توسط گاو مصرف می شوند، اسید اولئیک (بالا)، اسید لینولئیک (میانه) و الفا لینولنیک اسید (پایین).
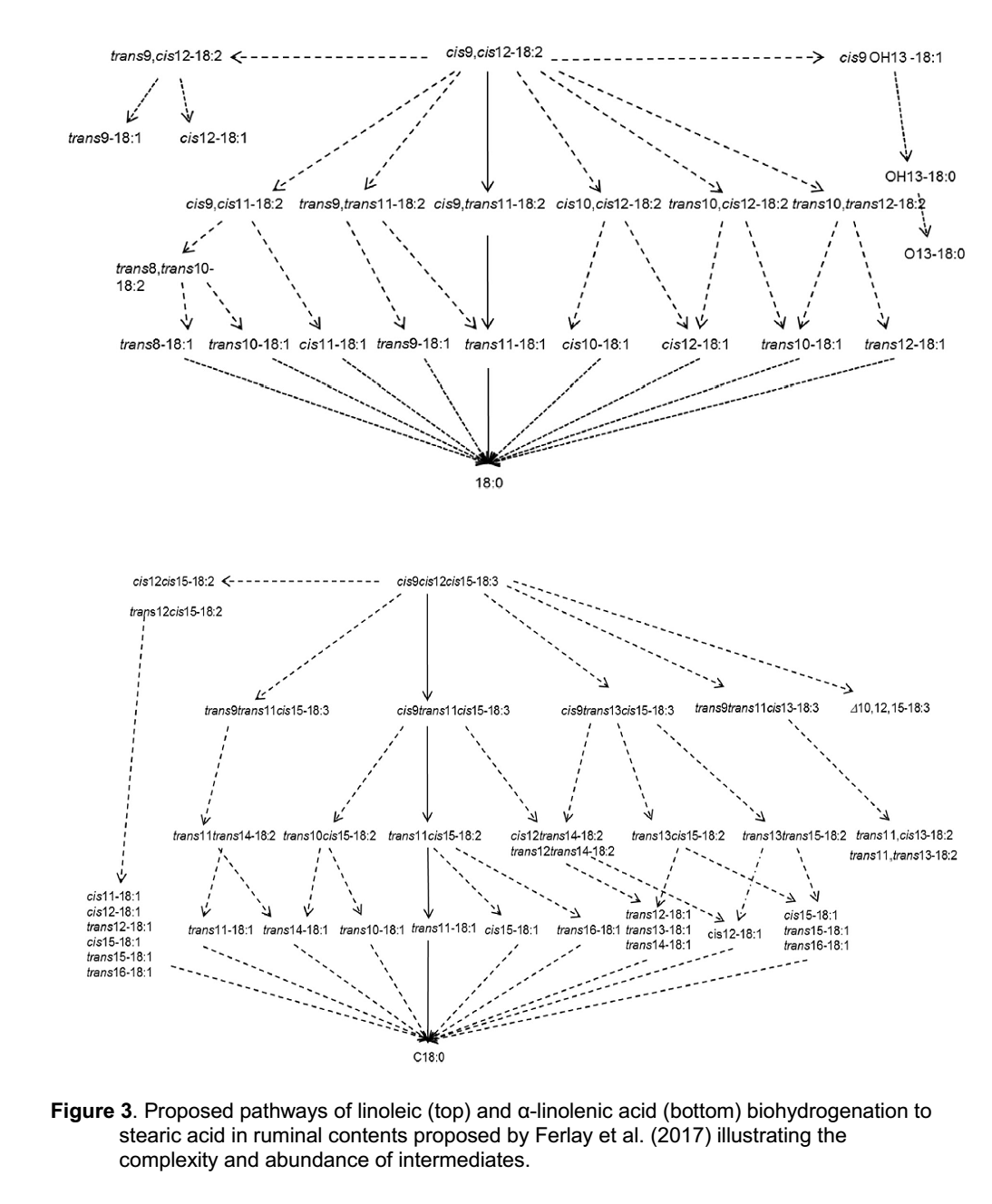
شکل 3. مسیرهای پیشنهادی بیوهیدروژناسیون لینولئیک (بالا) و آلفا لینولنیک اسید (پایین) به اسید استئاریک در شکمبه پیشنهاد شده توسطFerlay et al. (2017) که پیچیدگی و فراوانی واسطه ها را نشان می دهد.
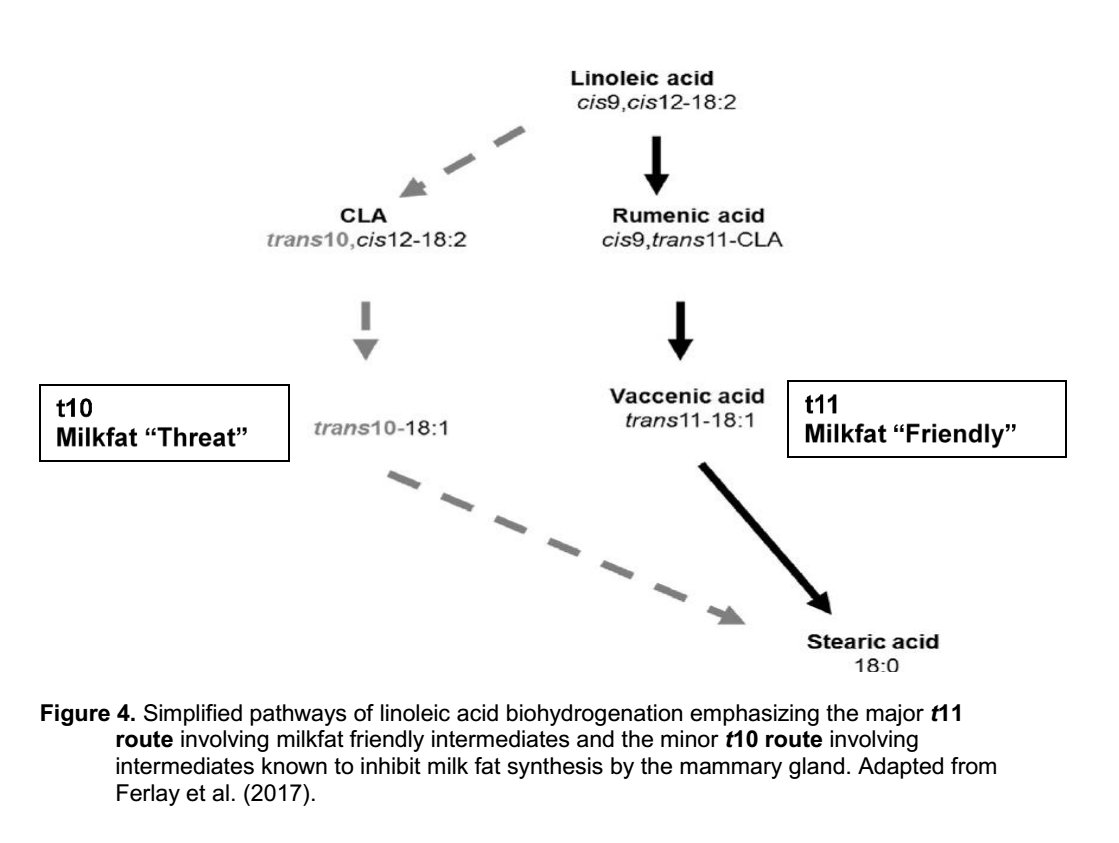
شکل 4. مسیرهای ساده بیوهیدروژناسیون اسید لینولئیک با تأکید بر مسیر اصلی t11 شامل واسطه های دوستانه چربی شیر و مسیر فرعی t10 شامل واسطه هایی که برای مهار تولید چربی شیر توسط غده پستانی شناخته می شوند برگرفته از Ferlay et al. (2017).
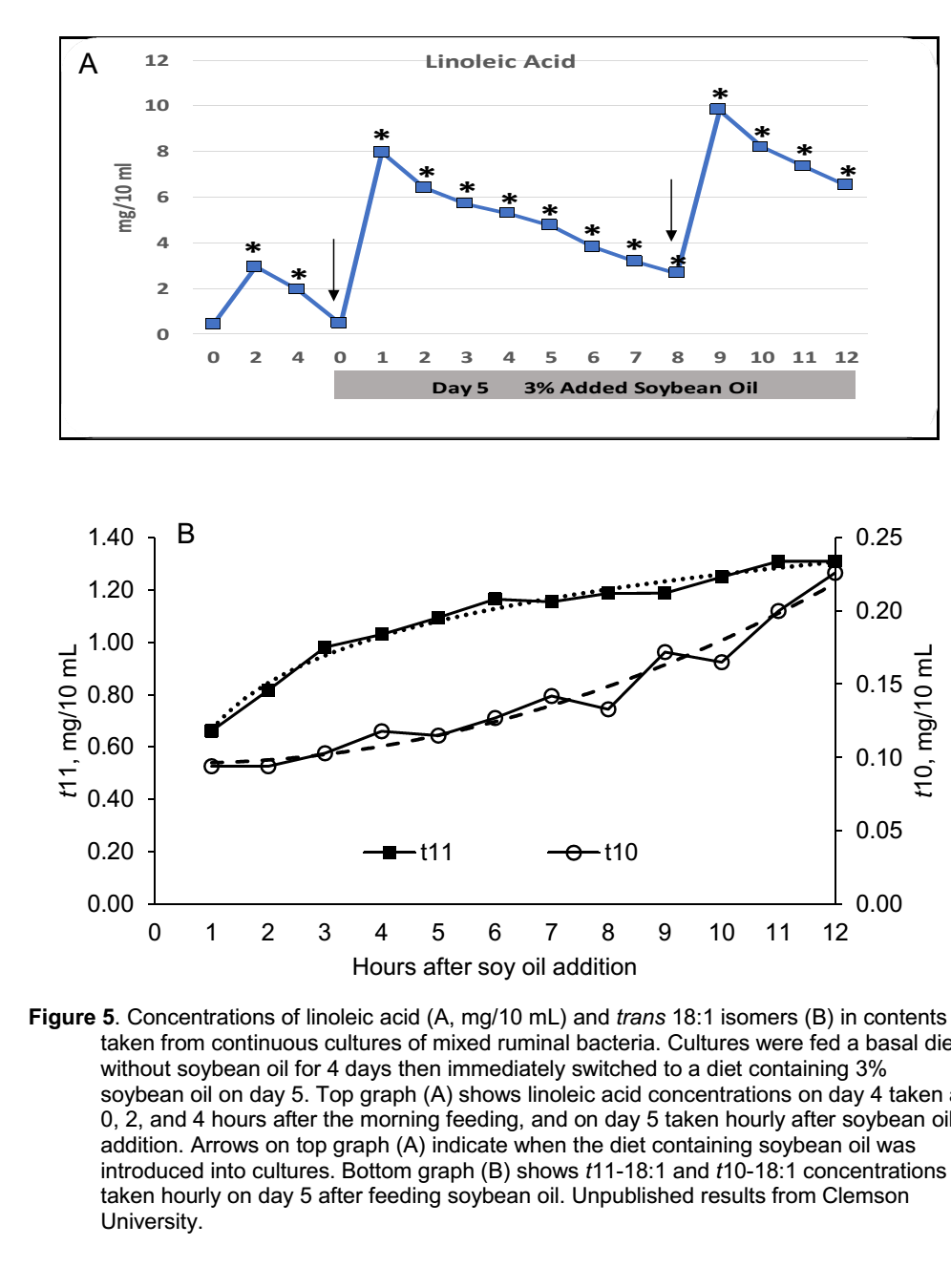
شکل 5. غلظت اسید لینولئیک (A، میلی گرم در 10 میلی لیتر) و ایزومرهای ترانس 18:1 (B) در محتویات گرفته شده از کشت پیوسته باکتری های مخلوط شکمبه. در محیط کشت به مدت 4 روز از یک جیره پایه بدون روغن سویا استفاده شد و بلافاصله به رژیم غذایی حاوی 3 درصد روغن سویا در روز 5 استفاده شد. شکل بالا(A) غلظت اسید لینولئیک را در روز 4 در 0 ، 2 و 4 ساعت بعد از تغذیه صبح نشان می دهد و در روز 5 هر ساعت بعد از افزودن روغن سویا. فلش ها در نمودار بالا (A) زمان افزودن روغن سویا به محیط کشت را نشان داده است. نمودار پایین (B) غلظت های t11-18: 1 و t10-18: 1 را نشان می دهد که روز 5 بعد از تغذیه روغن سویا به صورت ساعتی در روز اندازه گرفته می شود.
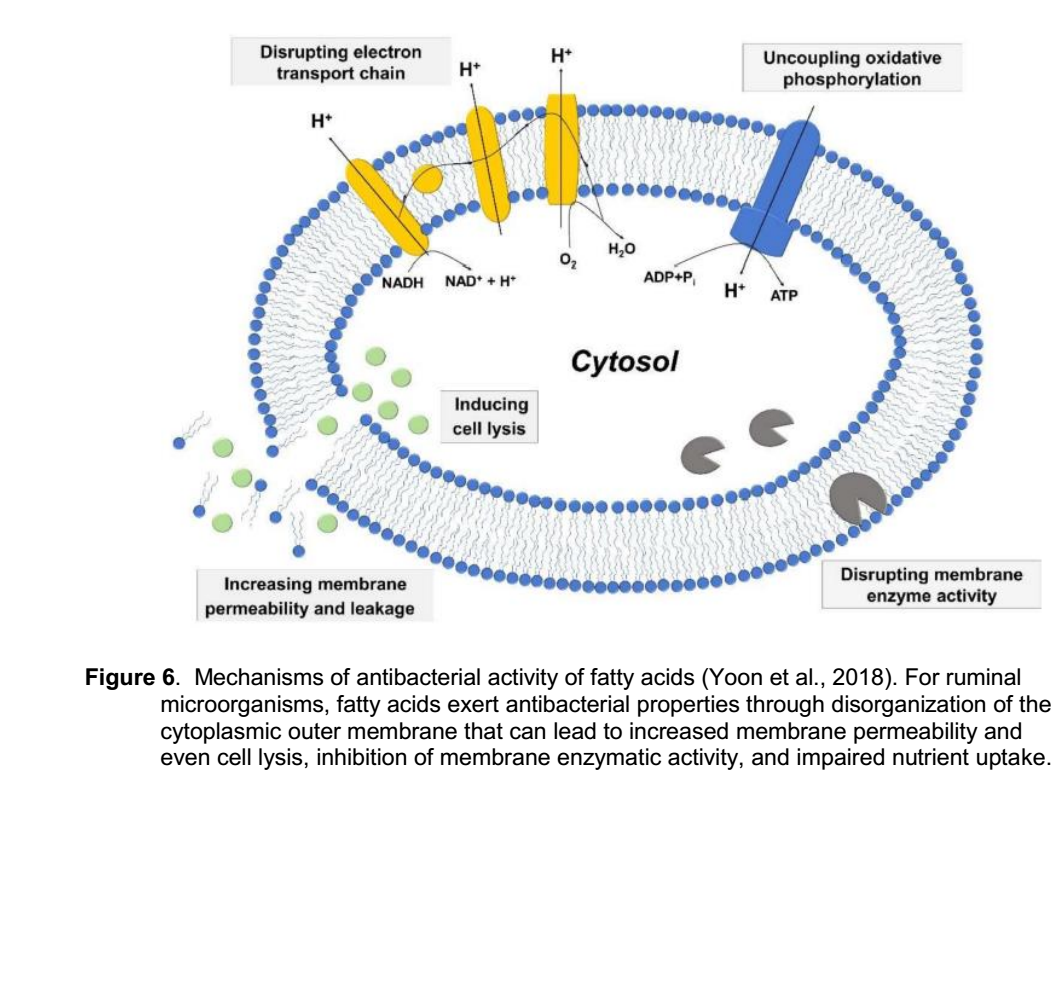
شکل 6. مکانیسم های فعالیت ضد باکتریایی اسیدهای چرب (Yoon et al., 2018). برای میکروارگانیسم های شکمبه، اسیدهای چرب از طریق ایجاد اختلال در غشای خارجی سیتوپلاسمی خاصیت ضد باکتریایی دارند که می تواند منجر به افزایش نفوذپذیری غشا و حتی تخریب سلول، مهار فعالیت آنزیمی غشا و اختلال در جذب مواد مغذی شود.
گروه علمی شرکت اگرین تک